Евкаріоти в пошуках свого коріння
23 Липня 2015 | Молекулярно про Рак
Категорія: Біологія
Теги: археї, архебактерії, бактерії, евкаріогенез, евкаріоти, еволюція, еукаріоти, мітохондрія, прокаріоти
Наприкінці травня 2015 року по стрічках новин різноманітних науко-орієнтованих інформаційних ресурсів пробігла блискавка, епіцентром якої став Університет Уппсали в Швеції. Вони замайоріли заголовками, які по суті можна звести приблизно до такого: “Нарешті знайдено втрачену еволюційну ланку між прокаріотами та евкаріотами!” І як часто буває в дослідженнях подібного плану, цей прорив став сукупним результатом багатьох років наполегливої праці та щасливої випадковості. Тож, що собою являє ця “втрачена ланка” і як вона вплинула на погляди вчених щодо еволюції евкаріот на нашій планеті?
Еволюція еволюції евкаріот
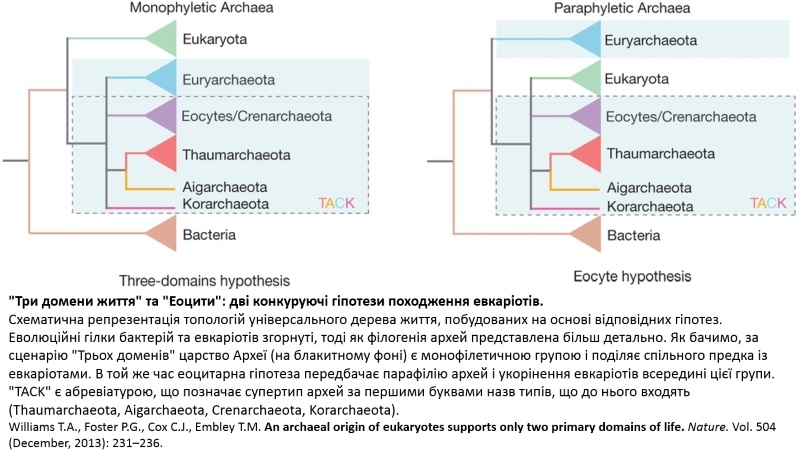
Згідно із хрестоматійним баченням еволюції життя на Землі, вперше сформульованим Карлом Воузом в 1977 році, на основі послідовностей 16S рибосомальної РНК та генів, задіяних в трансляції протеїнів, клітинні форми життя розподілено на три основні домени: прокаріоти у формі (1) бактерій та (2) архей і (3) евкаріоти. Виокремлення трьох доменів відображає наші наявні знання про особливості будови і функціонування представників відповідних груп організмів, але разом із тим ставить цілком природне питання: яким чином зазначені домени еволюційно пов’язані між собою? На цей рахунок від початку існувало дві основні гіпотези. Перша передбачала раннє відокремлення еволюційної гілки, що веде до евкаріот. Справді, в об’єднанні бактерій та архей за ключовою відмінною від евкаріот ознакою — відсутністю ядра — є певна логіка. Проте, з часом така гіпотеза виявилася нежиттєздатною. По-перше, вона не знайшла підтримки в царині палеонтології. По-друге, поступове зростання кількості геномів, для яких встановлена первинна послідовність нуклеотидів (секвенованих геномів), і паралельне покращення методів їх порівняння (філогенетичного аналізу) схилили шальки терезів на користь сценарію, в якому евкаріоти ближче споріднені з археями та ймовірно поділяють спільного предка, тоді як бактерії відгалужуються набагато раніше в ході еволюції. Таке бачення панувало впродовж тривалого часу, і всі висновки з наукових даних робилися на його основі. Але, як часто це буває із будь-якою динамічною науковою проблемою, нові результати, які не зовсім узгоджувалися із базовою концепцією, не забарилися.
Причиною цього насамперед стало збільшення кількості доступних геномів архей внаслідок значного поступу, власне, методів секвенування та комп’ютерної обробки первинних послідовностей нуклеотидів. Тут головно йдеться про методи так званого метагеномного аналізу. Їх застосування дозволило вирішити одну з найзначніших проблем мікробіології, знану як “некультивовані форми життя”. Більшість відомих нині архей складно виростити штучно, оскільки вони живуть в дуже тонко збалансованих екосистемах, унікально пристосованих для екстремальних умов, які просто неможливо відтворити в лабораторії. Разом із тим до недавнього часу усі методи секвенування передбачали виділення ДНК виключно з чистої культури клітин in vitro. Але на щастя сьогодні вже існують методи визначення окремих нуклеотидних послідовностей з суміші мікроорганізмів, отриманих з природних джерел, оминаючи необхідність культивування.
Отже застосування методів метагеноміки дозволило ідентифікувати велику кількість нових архей. Після детального аналізу якісного складу їхніх геномів стало зрозуміло, що багато з них містять послідовності, явно прямо споріднені із евкаріотичними. Нові дані кардинально змінили топологію універсального еволюційного дерева: одні археї наблизилися до евкаріот, інші віддалилися, і у підсумку евкаріотична гілка еволюції опинилася у щільному оточенні архей. Тобто все скидалося на те, що предка евкаріот слід шукати саме серед архей. Цю ідею ще у 1984 році сформулював Джеймс Лейк на підставі приголомшливої подібності структури рибосом Кренархей та сучасних евкаріот. Оскільки на той час цей тип архей звався “еоцити”, відповідно така гіпотеза походження евкаріот отримала назву “еоцитарна“. На жаль, самої лише структурної подібності, відомої на той момент, було недостатньо для повного обґрунтування зазначеної гіпотези, тому довгий час вона залишалася лише припущенням. І от на початку нового тисячоліття, після майже 30-річного безроздільного панування, концепція трьох доменів життя похитнулася — виявилося, що реально існують лише два домени — бактерії та археї — а евкаріоти походять від останніх. На цьому моменті, коли все нарешті стало зрозуміло, було б логічним перейти безпосередньо до неймовірного відкриття шведських вчених, проте є певна ускладнювальна обставина, яку просто-таки не можна оминути увагою: в евкаріот є мітохондрія.
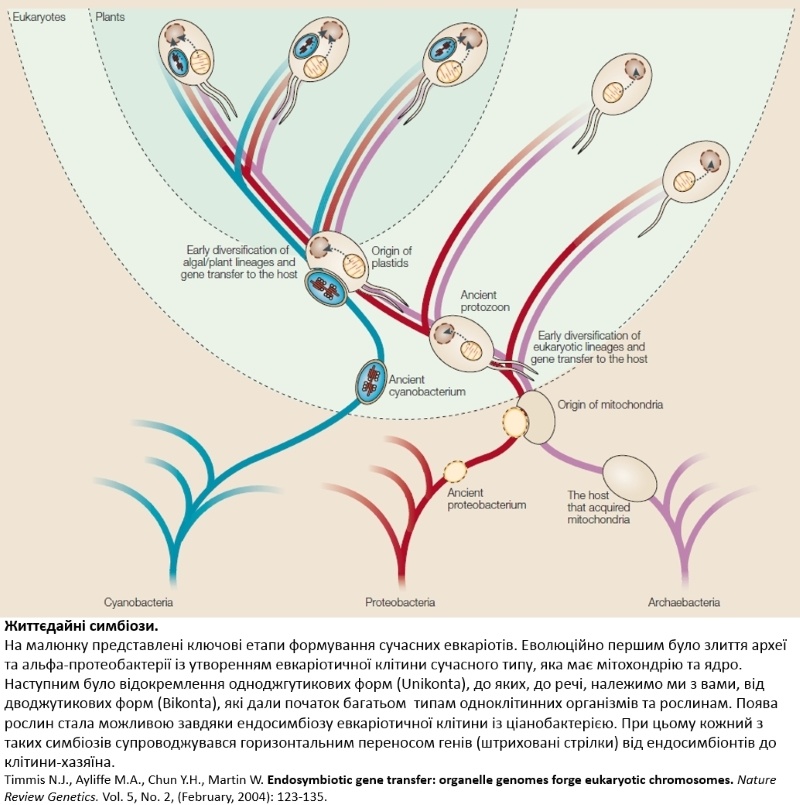
Найвизначніше злиття за всю історію життя
Сказати лише, що евкаріотам дісталися гени у спадок від архей — це сказали напівправду. Типовий евкаріотичний геном є в кращому випадку трикомпонентною химерою. По-перше, він містить, власне, архейні гени, що походять від прабатьківської клітини; по-друге — бактеріальні гени, що вказує на тісний симбіоз між археєю та альфа-протеобактерією, яка згодом перетворилася на мітохондрію (в рослин до цього додаються також ціанобактеріальні гени від іншого відомого симбіозу, який завершився утворенням хлоропласту); по-третє — унікальні гени, що стали результатом свого власного еволюційного шляху евкаріот і зумовили появу усього багатоманіття форм життя, які ми можемо бачити навколо себе неозброєним оком. Яким саме чином всі ці різні за походженням компоненти опинилися “під одним дахом” — це тема для іншої дискусії. Але які факти дають нам підставу стверджувати, що саме архея була батьківською клітиною, а альфа-протеобактерія була в ній “квартирантом” (виражаючись науковою лексикою — ендосимбіонтом), а не навпаки? Відповідь на це питання криється у тому, які функції виконують гени відповідного походження. Евкаріотинчі гени умовно поділяють на керівні (informational genes), які відповідають за фундаментальні процеси передачі інформації в клітині (реплікація, транскрипція, трансляція, ключові регуляторні каскади), і виконавчі (operational genes), що кодують ферменти різноманітних метаболічних шляхів. Так от виявляється, що гени архебактеріального походження здебільшого є керівними, тоді як виконавчі гени з більшою вірогідністю є бактеріальними за походженням. Такий невипадковий розподіл функцій свідчить на користь того, що ядерна ДНК бере свій початок саме від архей, а бактеріальні гени потрапили до евкаріотичного геному шляхом горизонтального переносу від ендосимбіонта.
Та незалежно від того, чиї гени є основними, злиття археї та бактерії — ключовий етап евкаріогенезу. Об’єднання потужностей двох принципово різних типів прокаріотичних організмів в одній клітині стало еволюційним поштовхом неймовірної сили, відгомін якого ми бачимо і сьогодні. З майже чотирьох мільярдів років існування життя на Землі, евкаріотичні форми населяють її лише протягом 1,2 млрд років, тож на утворення стійкого симбіозу між археєю та альфа-протеобактерією пішло чимало часу! Тобто такий симбіоз є лімітуючою стадією евкаріогенезу, і надзвичайна складність і стохастичність процесу його становлення пояснює чому евкаріотична клітина утворилася лише один раз за натуральну історію Землі. Розуміючи красу і важливість цього видатного злиття, вчені сформулювали концепцію “кільця життя”, де еволюційна гілка евкаріот є таким собі анастомозом двох прокаріотичних гілок. Ця ідея певною мірою повертає евкаріотам право називатися “третім доменом”, щоправда з оговіркою, що вони є композитною формою життя.
Але що стало передумовою злиття двох окремих організмів у один? Міркуючи над цим питанням, науковці натрапили на певне протиріччя. З одного боку складно побудована клітина вимагає достатньо об’ємного геному, щоб закодувати усі складові частини такої клітини. А підтримання великого за обсягом геному — це дорога розкіш з енергетичної точки зору. Тож, якщо клітина “бажає ускладнюватися”, вона від початку повинна мати деяку органелу для ефективного отримання енергії, таку як (ви вгадали!) мітохондрія. З іншого боку, щоб захопити і привласнити собі альфа-протеобактерію, архея має бути здатною до фагоцитозу. За цим коротким і простим не перший погляд біологічним терміном стоїть купа прогресивних цитологічних ознак, як-от добре розвинені актиново-міозинові філаменти, системи ремоделювання цитоплазматичної мембрани та ендосомального трафіку — ознаки, які завжди вважалися “монополією” евкаріот і якісно відрізняли їх від прокаріотів. Отже, дискусія про походження евкаріот, як і будь-яка гарна еволюційна проблема, врешті-решт зводиться до добре усім відомої форми “курка або яйце?”.
Витівка Локі чи дивовижне відкриття?
Розв’язати цю проблему — значить відшукати або принаймні реконструювати безпосереднього архебактеріального предка евкаріот. Для цього вчені вдаються до пошуку нових видів архей із різноманітних природних біотопів і в подальшому аналізують їхні геноми, шукаючи в них евкаріотичні “пра-гени”, для яких, до речі, навіть придумали свій термін із модним нині суфіксом “ом” — евкаріом. Такий підхід безумовно є перспективним (якщо не сказати, безальтернативним), оскільки він вже дозволив значно звузити коло пошуку ймовірних пращурів евкаріот до групи архей, що належать до супертипу під назвою TACK. Однак, на цьому фронті не без проблем. На сьогодні вдалося описати цілу низку архей, які мають ті чи інші характерні евкаріотичні ознаки (як-от фактори елонгації трансляції, актини, гістони, тощо). Проте усі вони мозаїчно розподілені серед різних представників згаданої групи.
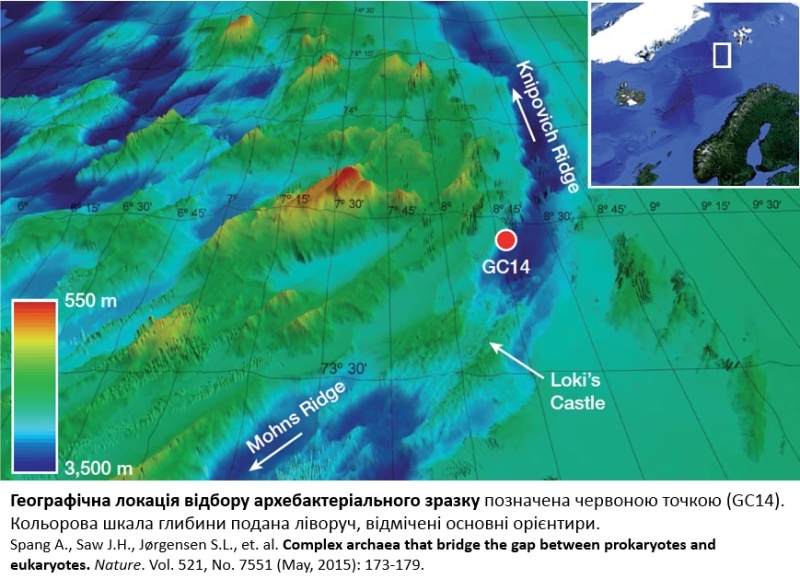
До останнього часу вченим не вдавалося знайти єдиної археї, яка б сполучала в собі всі ті прогресивні ознаки. У цього феномену навіть з’явилася своя назва “розпорошений евкаріом” (scattered eukaryome). Такий стан речей найімовірніше пояснюється тим, що архея-попередник із повним евкаріомом все-таки існувала (чи існує досі), а розпорошення відбулося шляхом диференційної втрати тих чи інших генів у ході редуктивної еволюції, що супроводжувала дивергенцію архей. І от в результаті довготривалої пошукової та аналітичної роботи шведськими вченими був знайдений мікроорганізм гідний бути кандидатом на роль універсальної археї-предка. Під час сканування проби, відібраної роботом на глибині майже три з половиною кілометри в Північному Льодовиковому Океані між норвезьким узбережжям та архіпелагом Шпіцберген, був охарактеризований новий тип архей із безпрецедентним набором генів, подібних до евкаріотичних. Місце взяття проби знаходилося в безпосередній близькості до гідротермального курця Замок Локі (Loki’s Castle), названого на честь скандинавського божества бешкету і обману. Відповідно, новий тип назвали Lokiarhaeota. Спершу було здійснено метагеномний аналіз зразка, і після обробки даних секвенування із застосуванням складних алгоритмів фільтрування і об’єднання даних, було зібрано майже повний (92 %) геном Локіархеї. Досліджуючи якісний склад її геному, вчені виявили разючу подібність локіархеальних та евкаріотичних актинів, а також знайшли ймовірні аналоги білків, що залучені в забезпеченні динаміки актинових філаментів (останні, до речі, описані у прокаріотів вперше). Іншою несподіваною знахідкою стала кількість генів, як кодують ГТФази надродини Ras — важливі сигнальні молекули, що контролюють перебудови цитоскелету, утворення мембранних везикул, транзит макромолекул між ядром і цитоплазмою в евкаріотичних клітинах. Парадокс ситуації полягає в тому, що кількість генів ГТФаз локіархей перевищує таку для деяких евкаріот, тоді як до цього моменту серед прокаріотичних організмів траплялися лише поодинокі аналоги ГТФаз. Такі особливості клітинної мікроорганізації характерні, наприклад, для амеб, що здатні до активного захоплення здобичі. Звичайно, зі самої лише генної композиції важко робити висновки щодо реальних можливостей даної археї. Та все одно вчені зауважують, що здобуття мітохондрії скоріше всього відбулося вже після того, як почався процес набуття археєю ознак складно побудованої клітини. На додачу, дослідники також виявили гени, чиї продукти відповідають за ендосомальний трафік та убіквітин-залежну деградацію протеїнів, завершуючи таким чином збирання розпорошеного евкаріому.
Опоненти і скептики зазначили б, що такі екстраординарні результати можна пояснити забрудненням архебактеріального зразку сучасними евкаріотами. Але вчені наголошують на абсолютній безпомилковості своїх висновків з метагеномних даних. По-перше, ПЛР із застосуванням специфічних праймерів не виявила слідів евкаріотичної 18S рибосомної РНК, по-друге, висока якість збірки геному засвідчила, що пра-евкаріотичні гени обрамлені типовими архебактеріальними генами і організовані у вигляді характерних для прокаріотів оперонів. Отже все скидається на те, що це не витівка бешкетника Локі (виражаючись науковою лексикою — не артефакт), а дійсно серйозне відкриття в дослідженні еволюційної історії евкаріот. Зрештою, біологи-еволюціоністи — це лишень групка евкаріотичних організмів, які прагнуть відшукати своє коріння.
Даниїл Грибчук для Моєї Науки
Дні науки

Наші проєкти

Щеплення Правдою



Обговорення