Гігантські віруси: втрачена гілка на дереві життя або молекулярні клептомани?
23 Травня 2017 | Молекулярно про Рак
Категорія: Біологія
Теги: віросвіт, вірусологія, гігантські віруси, дерево життя, еволюція, молекулярна біологія, трансляція
Гігантські віруси є живим втіленням поняття “загадка природи”. Розміри їхніх віріонів щонайменше в п’ять разів перевершують такі у решти вірусів. Наприклад, розмір вірусної частки Пандоравірусу досягає одного мікрометру, тому його нескладно побачити у звичайний мікроскоп (через це його спершу взагалі переплутали із бактерією). Гігантські віруси інфікують одноклітинні організми. Найбільше їх знайдено серед амеб (Mimiviridae, Pandoravirus, Marseillevirus та Pithovirus), але також вони відомі у різноманітних водоростей (Phycodnaviridae). Відкриття гігантських вірусів на початку ХХІ століття заскочило зненацька найвідоміших у світі вчених, змусивши одних висувати неймовірні теорії їх походження, а інших — ламати голову над їх спростуванням.
В одній зі своїх нещодавніх статей я писав, що гігантські віруси “вартують окремого розгляду”. І зараз огляд на цю тему є як ніколи своєчасним, оскільки нещодавно (7-го квітня 2017) в журналі Science було опубліковано повідомлення про новий раніше небачений гігантський вірус названий Klosneuvirus (KNV) на честь невеличкого містечка Клоштернойбург в Австрії. Виняткова важливість цієї знахідки полягає не в самому факті опису нового вірусу (насправді, цей вірус ніхто навіть не виділяв в чисту культуру, щоб ретельно описати!), а в тому, які білки закодовані в геномі цього вірусу. Виявилося, що KNV має майже повний набір з дев’ятнадцяти аміноацил тРНК синтетаз (aaRS) — ключових ензимів для вбудовування двадцяти типів амінокислот в процесі біосинтезу усіх протеїнів (трансляції). До цього рекорд за кількістю aaRS ензимів (7 штук) належав іншому гігантському вірусу Megavirus chilensis (MCV), що належить до роду Mimivirus. Краса ситуації полягає в тому, що трансляція є процесом характерним лише для клітинних організмів, а віруси за визначенням є молекулярними паразитами, які користуються послугами (тобто, нахабно відбирають) трансляційного апарату представників одного з трьох доменів життя — архей, бактерій чи еукаріотів. Тож, сім вирусних aaRS синтетаз можна вважати цікавим винятком з правил. Але ж вірус, в якого є aaRS синтетази аж до дев’ятнадцяти амінокислот — це щось абсолютно нечуване! Через це Klosneuvirus являє собою унікальну можливість перевірити гіпотезу приналежності гігантських вірусів до містичного четвертого домену життя шляхом порівняння вірусних та клітинніх аміноацил тРНК синтетаз.
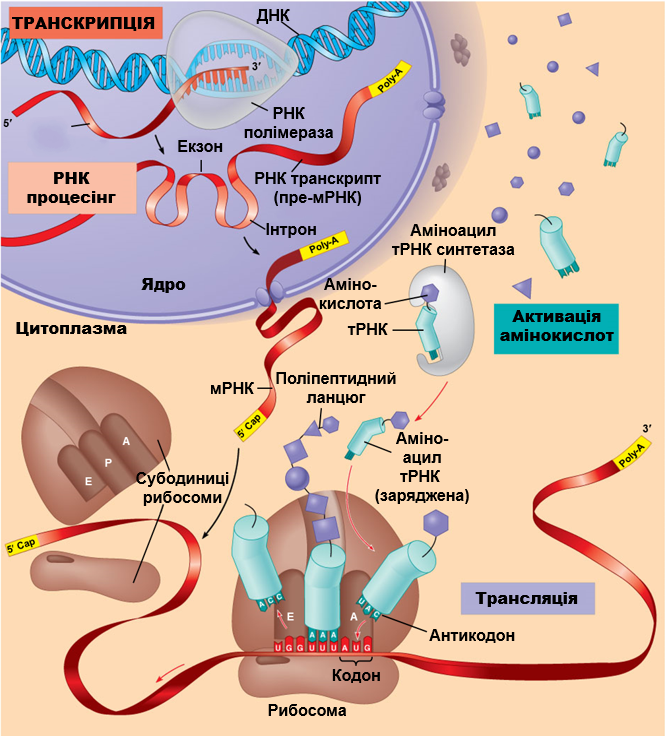
Але про все по черзі: як завжди, спочатку трошки контексту, а далі — детально розберемо, які висновки було зроблено із вищезгаданого дослідження.
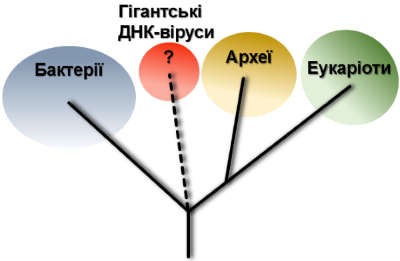
Усі живі організми традиційно поділяють на три великі групи, що звуться “доменами життя”. Основою для такого поділу слугують відмінності в найбільш важливій молекулярній машині клітини, що відповідає за побудову білків — рибосомі. Відтак, рибосоми еукаріотів, архей та бактерій різняться між собою за структурою і кількістю елементів. В свою чергу, віруси, так само традиційно, відносять до неклітинних форм життя, оскільки вони не кодують жодних компонентів біосинтезу білків. Це принципове обмеження робить неможливим з’ясування положення вірусів на дереві життя. Міркування над цією проблемою з рештою призвело до консенсусного рішення про те, що не існує єдиної точки виникнення вірусів. Натомість, різні віруси виникали незалежно на різних етапах розвитку життя на Землі. Але це ніяким чином не пояснювало сам механізм утворення вірусу. На цей рахунок існує щонайменше три гіпотези, одну з яких ми якраз-таки обговоримо в світлі сучасних досліджень гігантських вірусів. Це теорія регресивної еволюції або дегенерації, згідно з якою віруси в минулому були клітинами, що втратили велику частину клітинних систем включно з компонентами трансляції білків. Можливою причиною такого радикального спрощення вчені називають паразитичний спосіб життя: поступово гіпотетичний предок вірусів ставав усе більш залежним від клітини-хазяїна, втрачаючи свої частини, що ставали непотрібними. Довгий час було абсолютно незрозумілим яким чином клітинний організм може дійти до стану вірусу, оскільки навіть найпростіший мікроб мав набагато складнішу будову і більшу кількість і різноманітність генів, ніж будь-який об’єкт класичної вірусології. Але на початку 2000-х гіпотеза регресії несподівано отримала потужний поштовх з появою перших повідомлень про гігантські віруси (ГВ) — вчені розгледіли в них імовірні перехідні форми між “складними” клітинами і “простими” вірусами. Екстраординарні знахідки як правило провокують екстраординарні пояснення. Проте, щоб залишатися неупередженим, варто відмітити, що в той самий час розроблялася альтернатівна гіпотеза щодо походження ГВ. Вона ґрунтувалася на діаметрально протилежному твердженні: гігантські віруси пішли від менших ДНК-вірусів шляхом поступового накопичення додаткових генів. І як зазвичай буває в хорошому науковому блокбастері, в обох пояснень були сильні і слабкі аргументи. Тож, спробуємо в цьому розібратися.
Основна складність у вивченні гігантських вірусів полягає в тому, що вони є вкрай нетиповими, в буквальному сенсі “рвуть шаблон”, руйнують усталену схему класифікації живого. Це випливає з аналізу якісного та кількісного складу геномів ГВ: вони почасти містять гени, характерні лише для клітинних організмів, а також за розмірами вони входять в діапазон останніх. Відтак ГВ розмивають межі між клітинними формами життя та вірусами.
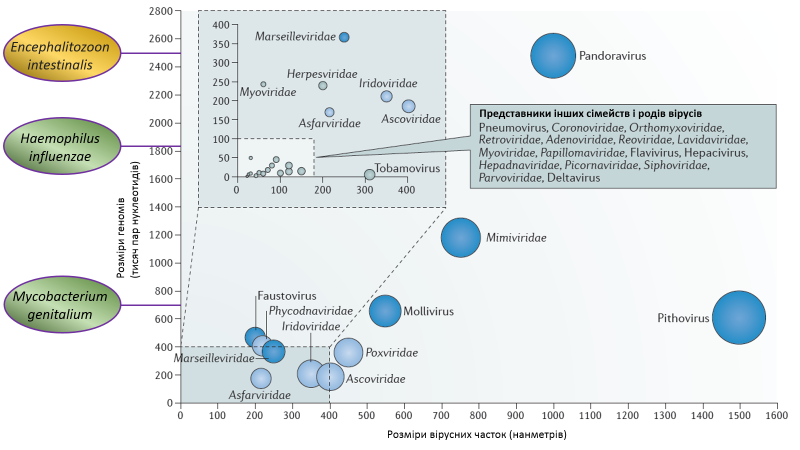
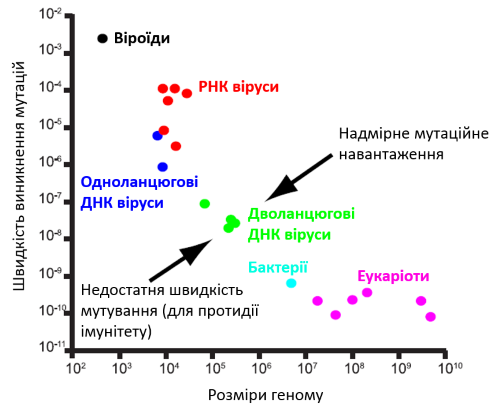
Донедавна вважалося, що розміри геному ДНК вірусу не можуть перевищити позначку в півмільйона пар нуклеотидів, оскільки на нього діють принаймні два еволюційні обмеження. По-перше, чим більший геном, тим більш складним і тривалим є процес його відтворення, а для вірусу питання швидкості є питанням виживання. По-друге, ензим, який виконує копіювання геному — ДНК-полімераза —, схильна робити помилки. Насправді, неточність — це фундаментальна характеристика всіх без винятку полімераз (в клітинних організмах так само). Але вирусні полімерази припускаються значно більшої кількості огріхів. З одного боку, це дозволяє їм швидше еволюціонувати і протидіяти імунітету. Та з іншого боку це збільшує ризики втратити функціонально важливий ген, що вестиме до нежиттєздатності вірусу. Вважається, що для малих та середніх вірусів це не становить серйозної загрози через високу репродуктивну активність (віруси беруть кількістю, а не якістю). Проте, із зростанням величини геному і числа життєво-важливих генів, частота критичних поломок непропорційно збільшується. Поява на радарі сучасної науки гігантських вірусів чітко вказувала на явні прогалини в нашому розумінні факторів та напрямків еволюції ДНК вірусів в цілому.
Отже, кричуща невідповідність між теорією та спостереженнями вимагала пояснень. І перші зачіпки науковці отримали аналізуючи якісний склад генномів мімівірусів групи A, B та C. Як ви вже здогадалися, зараз мова піде про універсальні консервативні гени необхідні для синтезу протеїнів: аміноацил-тРНК синтетази (aaRS). Як зазначалося вище вони є характерними лише для клітинних організмів, тож їх наявність у вірусів сама собою була сенсацією. Але ще більший подив у науковців викликали результати аналізу походження цих генів. Виявилося, що більшість з них (окрім aaRS для тирозину, TyrRS) утворювали окрему еволюційну гілку на дереві життя відмінну від усіх інших живих істот. Тобто віруси мали власні ні на що не схожі гени для забезпечення одного з основних процесів, на якому базується життя, — трансляції! Логічним висновком із цього стало те, що гігантські віруси є четвертим рівноправним членом елітного клубу «Домени Життя» поряд із бактеріями, археями та еукаріотами. Щодо тирозинової аміноацил тРНК синтезази, вона була явно споріднена із такою у амеби — природного хазяїна мімівірусів. Це вказувало на можливість горизонтального переносу TyrRS гену від амеби до вірусу. Проте, з тією ж вірогідністю процес міг пройти і в зворотному напрямку. Розв’язати це питання на той час було неможливо через невизначене філогенетичне положення самої амеби (йдеться про Entamoeba histolytica) на еукаріотичному еволюційному дереві.
Інший цікавий висновок випливав з аналізу розподілу aaRS генів серед представників трьох різних груп мімівірусів. Відтак, у групах A та B спостерігається відсутність певних генів аміноацил тРНК синтетаз, тоді як в групі C присутні усі aaRS, характерні для груп A та B, плюс одна унікальна. Це було свідченням того, що спільний предок мімівірусів мав значно ширший репертуар aaRS генів, і, відповідно, представники групи C є найбільш схожими на предкову форму. Надалі різні еволюційні лінії мімівірусів незалежно одна від одної втрачали ці гени в ході пристосовування до конкретних хазяїв, що є наочним прикладом регресивної еволюції. Екстраполювавши цю тенденцію, дослідники прийшли до висновку, що спільний предок мімівірусів мав будову значно складнішу, ніж всі сучасні мімівіруси, і ймовірно міг бути клітинним організмом.
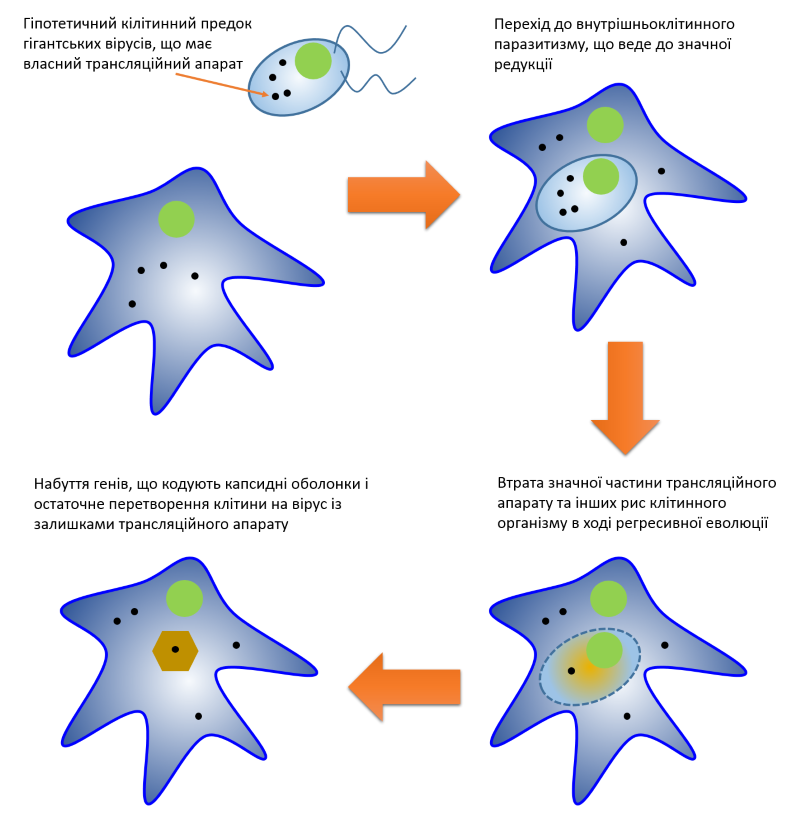
Та попри всю красу та проривність теорії, в ній залишалося чимало недоліків: невідповідність філогенії TyrRS був лише одним з багатьох. І поки захисники чотирьох доменів старанно відповідали на всі закиди опонентів, інші готували фінальний удар по теорії.
Першим дзвоником стала демонстрація генетичної спорідненості всіх ядерно-цитоплазматичних великих ДНК вірусів (nucleo-cytoplasmic large DNA viruses, NCLDVs). Ця група, окрім гігантських вірусів на кшталт мімівірусу та пандоравірусу, включає також менші віруси такі як вірус віспи (наразі запропоновано об’єднати всі родини NCLDVs — Poxviridae, Asfarviridae, Iridoviridae, Ascoviridae, Mimiviridae, Phycodnaviridae та Marseillevirus — в порядок Megavirales). Такі результати випливали з аналізу п’яти універсальних консервативних генів присутніх в усіх цих вірусів. Отже, з цього моменту еволюцію мімівірусів слід було розглядати лише в контексті наявних даних про NCLDVs віруси, а не окремо як це робилося раніше. У ході порівняння мімівірусів із їхніми NCLDV-побратимами виявилося, що і величезний розмір геному (більше мільйона пар нуклеотидів) і наявність генів, задіяних в трансляції протеїнів (зокрема aaRS), є радше винятковою особливістю мімівірусів, ніж загальною закономірністю організації представників зазначеної групи. Противники теорії чотирьох доменів використали це як аргумент на користь більшої вірогідності сценарію, за яким і гігантські віруси пішли від менших шляхом запозичення генів у клітин. В той же час, поборники теорії наполягали, що решта вірусів NCLDVs групи були редукованою формою мімівірусів, які ще далі просунулися на шляху дегенерації.
Разом із тим, більш радикальні противники концепції чотирьох доменів взагалі відкидали саму можливість з’ясувати міру спорідненості гігантських вірусів і клітинних організмів, вказуючи та суттєву методологічну помилку такого аналізу. Як зазначалося вище, вірусна ДНК полімераза є менш точною, ніж клітинна, і відповідно ДНК віруси еволюціонують швидше за клітини. Тож, навіть якщо вірус отримав певний ген шляхом горизонтального переносу від клітини, то протягом відносно нетривалого часу послідовність такого гену зміниться до невпізнаваності, і клітину-донора вже неможливо буде встановити. Відповідно, на еволюційному дереві, побудованому за такими генами, віруси будуть гарантовано ставати у конфігурацію четвертого домену. В даному випадку розбіжності в послідовоностях досліджуваних генів будуть обумовлені не унікальністю еволюційної лінії вірусів, а лише швидшими темпами їхньої еволюції загалом.
На додачу до вищесказаного, детальний аналіз генів мімівірусу показав, що частина з них не є характерними для амеб, а походять від інших еукаріотів і навіть бактерій. Це спостереження зайвий раз вказує на схильність мімівірусів привласнювати чужі гени. Власне, приблизно з такими розкладами ми виходимо на останнє повідомлення із царини гігантських вірусів, яке, відповідно до поважних видань, «розпалює суперечки навколо [їх] походження з новою силою» (хоча, як на мене, то доречніше було б сказати: «заганяє останній цвях у віко труни теорії чотирьох доменів»).
https://gfycat.com/UnacceptableAcrobaticCoyote
Рис. 6. Захоплення нових генів ДНК вірусами. Джерело: Nature.
Отже, 7-го квітня 2017 року група вчених з Австрії та Сполучених Штатів публікує повідомлення про новий тип гігантського вірусу, знайдений в одній з цистерн водоочисної станції в містечку Клоштернойбург поблизу Відня. Вірусна ДНК в зразку води була виявлена шляхом так званого метагеномного секвенування — методу, що дозволяє встановлювати майже повні послідовності ДНК в матеріалі з довкілля без потреби лабораторного культивування. Додатково, дослідники також продемонстрували наявність вірусних часток (капсидів) за розміром та формою схожі на такі в гігантських вірусах. Довжина геному Клоснойвірусу (KNV) склала близько півтора мільйони пар нуклеотидів. Технологія метагеномного секвенування передбачає поступову збірку довгих послідовоностей із значно коротших (довжиною лише 100 нуклеотидів). Але вчені запевняють нас у високій якості зібраного геному, вказуючи на достатню глибину секвенування і надійне перекриття фрагментів, однорідність відносного вмісту GC пар та наявність вірус-специфічних генів по всій довжині геному. Побудова еволюційного дерева за п’ятьма універсальними консервативними генами, характерними для NCLDVs вірусів, чітко вказувала на спорідненість KNV із мімівірусами. Але водночас, KNV мав майже 2500 типів генів, раніше ніколи не бачених в мімівірусах. Співставлення наявності/відсутності генів із еволюційними траєкторіями мімі- та колоснеовірусів свідчило про значне переважання випадків набуття нових генів над їх втратами впродовж історичного розвитку (філогенезу) зазначених двох родів вірусів.
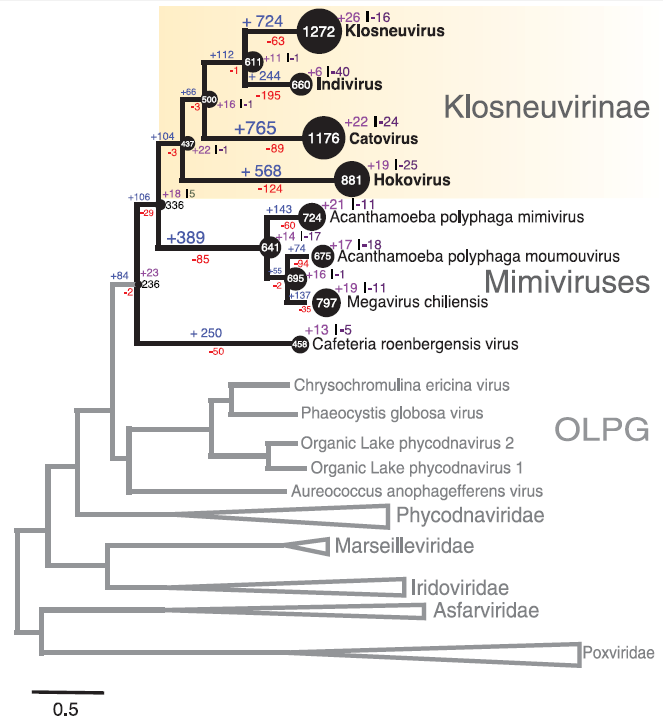
Але набагато показовішими є результати філогенетичного аналізу аміноацил-тРНК синтетаз KNV. Як зазначалося у вступі, KNV має 19 aaRS генів, що значно перевершує їхню кількість у мімівірусів. У разі, якщо гігантські віруси є реліктом епохи останнього універсального спільного предка (Last Universal Common Ancestor, LUCA), ми очікуємо, що усі ці гени повинні мати унікальну еволюційну історію нерозривно пов’язану із розвитком, власне, їх носіїв — вірусів. Натомість, за даними авторів лише один з 19-ти генів (аміноацил-тРНК синтеназа для ізолейцину) крок за кроком повторює еволюційний шлях гігантські вірусів. До пояснення цього спостереження зовсім не обов’язково залучати походження ДНК вірусів від ефемерної четвертої гілки життя. Простіше припустити, що відбулася єдина подія захоплення гену ізолейцинової aaRS спільним предком мімі- та клоснеовірусів. Щодо решти 18 aaRS генів, найбільш вірогідний сценарій передбачає їх незалежне набуття в різний час шляхом горизонтального переносу від різних клітинних організмів. Цікаво, що два інші aaRS гени (для гістидину та глутамінової кислоти) направду утворюють 4-й домен між бактеріями та археями, але дослідники пояснюють це швидшими темпами еволюції даних генів (див. вище). Загалом висновки зроблені за результатами дослідження є невтішними для прибічників теорії 4-го домену. Гігантські віруси більше нагадують кишенькових крадіїв генів ніж древню еволюційну гілку життя.
Такі результати видаються доволі логічними, оскільки гігантський вірус з майже повним набором aaRS генів все одно залишається по своїй суті вірусом і жодним чином не нагадує клітину. З іншого боку найбільш редукований організм відомий науці, який в минулому був клітиною — мітохондрія —, зовсім не схожа на вірус і анітрохи не втратила трансляційний апарат. Разом із тим, якщо дослідники дійсно мають намір переконати нас в тому, що гігантські віруси утворилися шляхом поступового накопичення генів, узятих від клітин, вони повинні бути готові відповісти на серйозне запитання: “Навіщо вірусам свої гени біосинтезу білку, в чому еволюційний сенс такої клептоманійної поведінки, так би мовити, адже решта вірусів повністю задовольняє свої потреби в експресії генів за рахунок клітинних факторів?”. Це питання вимагає глибинного розуміння біології мімівірусів, якого наразі немає. Поки що вчені втішаються лише спекуляціями. Можливо, наприклад, власні аміноацил тРНК синтетази потрібні вірусам для забезпечення безперебійного синтезу протеїнів у разі, якщо клітина-хазяїн вимикає процеси біосинтезу в рамках анти-вірусної відповіді. Інша гіпотеза бере до уваги той факт, що геноми вірусів та амеб різняться за відносним вмістом GC нуклеотидів ДНК (у амеб їх більше), тому вірус використовує трохи інші кодони для означення тих самих амінокислот. Тож, власні aaRS гени (а також гени транспортних РНК) потрібні вірусу для компенсації невідповідностей його генетичного коду із таким в клітині-хазяїні. В продовження цього міркування вчені зазначають, що чим більше вірус має генів для забезпечення синтезу протеїнів, тим менше він залежить від генетичного контексту, в якому опиняється, і тим ширше коло його потенційних жертв. Для вірусу, що спеціалізується на одноклітинних організмах, це неабияка вигода, оскільки в довкіллі буває важко знайти великі одноманітні популяції протистів певного виду, а відтак, доводиться бути менш перебірливим і готовим до всього. Перевірка цих гіпотез очевидно спонукатиме до подальшого вивчення гігантських вірусів, які ще приховують від нас величезну кількість таємниць.
Даниїл Грибчук для Моєї Науки
Джерела:
Schulz F., Yutin N., Ivanova N.N. et. al. (2017). Giant viruses with an expanded complement of translation system components. Science 356 (6333). c. 82-85. doi: 10.1126/science.aal4657.
Abrah?o J.S., Ara?jo R., Colson P., La Scola B. (2017). The analysis of translation-related gene set boosts debates around origin and evolution of mimiviruses. PLoS Genet. 13 (2). c. e1006532. doi: 10.1371/journal.pgen.1006532.
Leslie M. (2017). Giant viruses found in Austrian sewage fuel debate over potential fourth domain of life. Science. doi: 10.1126/science.aal1005.
Reardon S. (2017). Giant virus discovery sparks debate over tree of life. Nature News. doi: 10.1038/nature.2017.21798.
Colson P., La Scola B., Levasseur A., et. al. (2017). Mimivirus: leading the way in the discovery of giant viruses of amoebae. Nat Rev Microbiol. 15 (4). c. 243-254. doi: 10.1038/nrmicro.2016.197.
Holmes EC. (2011). What does virus evolution tell us about virus origins? J Virol. 85 (11). c. 5247-5251. doi: 10.1128/JVI.02203-10.
Дні науки

Наші проєкти

Щеплення Правдою



Обговорення