Втеча ембріона із зони, трохи тюремного фолькльору
10 Вересня 2019 | Olga Maliuta
Категорія: Біологія
Теги: біологія розвитку, ембріон, запліднення, хетчинг
Щодня в ембріологічній лабораторії я як ембріолог бачу велику кількість людських ембріонів на різних етапах розвитку з нульового по 5 день їх існування. Це звична справа – оцінювати їхню морфологію, слідкувати за тим, як іде розвиток, та давати прогнози, який з ембріонів краще використати для підсаджування в матку пацієнткам, які заморожувати, а які й утилізувати. І от один надзвичайно красивий і, я б сказала, «жирний» ембріон 5-го дня розвитку змусив мене задуматись про один момент в житті ембріона, який називається «хетчингом» (англ. hatching) – вилуплення ембріона з оболонки.
Zona pellucida, або блискуча оболонка, початково оточує ооцит. Це ніби шкаралупа яйця, лише в нас вона м’яка і гелеподібна, а не тверда і крихка. ЇЇ задача захищати ооцит та ембріон від механічного пошкодження та бактерій і вірусів. Окрім захисту вона вміє пропускати хороших хлопців – сперматозоїдів – до яйцеклітини на побачення, ну…одного бажано.

Блискуча оболонка складається з 4 білків глікопротеїнів, які називаються ZP1, ZP2, ZP3 i ZP4. Майже всі вони виробляються кумулюсними клітинами, що оточують яйцеклітину у фолікулі під час дозрівання і після овуляції, які як няньки доглядають, годують і підтримують її та трясуться над нею як над нерозбитим яйцем.
ZP1 працює як структурний білок. ZP2 допоміжний білок у зв’язуванні сперматозоїда. Після запліднення ооцита цей білок модифікується, у нього з’являється декілька дисульфідних зв’язків у зміненій структурі й він стає нездатний більше зв’язати й пропустити сперматозоїд. Це захищає від поліспермії, тобто проникнення кількох сперматозоїдів. ZP3 пропускає лише сперматозоїди свого виду і захищає від міжвидового схрещування. Тому віслюк з мультика Шрек може покохати дракониху, але народження драковіслючків маловірогідно. Цей же білок запускає акросомальну реакцію сперматозоїда. Є дані, що він синтезується в першій профазі мейозу безпосередньо в розбухшому ядрі ооцита. ZP4 теж допомагає активувати сперматозоїди, але з часом перестає працювати на зв’язування, що теж захищає від проникнення великої кількості сперматозоїдів. Четвертий білок є у нас, пацюків, хом’яків, кролів, а от у більшості ссавців його нема. І у мишей його нема. В гені цього білка у мишей велика кількість стоп кодонів і білок просто не синтезується.
Після того як сперматозоїд пробурив собі хід в оболонці й злився з мембраною ооцита, в ньому (в ооциті) запускається так звана кортикальна реакція (Рис.1). По-перше, змінюється електричний потенціал мембрани і вона не здатна більше зв’язати інший сперматозоїд. По-друге, в простір між оболонкою вивільняється вміст кортикальних гранул, що багаті на глікозаміни, наприклад, гіалуронову кислоту. Ці речовини блокують проникнення інших сперматозоїдів і ущільнюють Zona pellucida. (Детальніше про ці та інші процеси можна прочитати у статті “Про незбагненне 2. Злиття двох планет”. – Ред.)
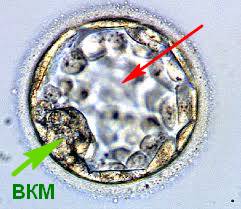
Далі йде швидкий і динамічний розвиток. Приблизно через 14 годин можна спостерігати 2 про-ядра зиготи – чоловічий і жіночий пронуклеуси. Після їхнього злиття ембріон починає дробитись. 48 годин – і це вже 4-клітинний ембріон. 72 години – 8 клітин. 96 годин – починається компактизація клітин і формується морула, що більше схожа на пережовану жувальну гумку. Вона настільки компактна, що не видно меж між клітинами. З часом в середині морули утворюється порожнина, яка швидко розширюється. На 120тій годині розвитку ембріон виглядає як кулька з порожниною – бластоцелем. Видно щільну групу клітин ВКМ – внутрішньоклітинну масу, що стане плодом і дитиною в майбутньому, та трофектодерму – розпластані клітини, що вистилають з середини всю блискучу оболонку, які дадуть початок плаценті. Це перша диференціація на тканини. Сам ембріон тепер носить назву «бластоциста» (Рис.2а).
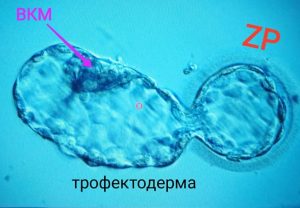
І ось тут виникає проблема. Ресурси, накопичені в яйцеклітині, вичерпуються, а місця в такій шкаралупці стає катастрофічно мало. Що повинна робити бластоциста? Тікати! Тікати з цієї в’язниці й зони. Бластоциста починає серію механічних рухів, вона періодично колапсує, стискається, зменшуючи об’єм бластоцелю. Після чого швидко накачує в порожнину більшу кількість рідини та розширюється і з ударним тиском розтягує оболонку. В неї немає ще ніяких м’язів, і вся механіка процесу- це лише накачування і відкачування рідини в порожнину. Зазвичай достатньо 3-4 циклів колапсування-розширення, щоб прорвати оболонку, та ще декілька для повного роздягання. На відео можна подивитись, як виглядає цей процес. У перехідний період ембріон виглядає як гантеля, коли частина бластоцисти уже вилупилась, а частина ще всередині (Рис.2б). Зазвичай прорив стається на полюсі протилежному до ВКМ, деякі вчені звернули на цей факт увагу і почали шукати причину.
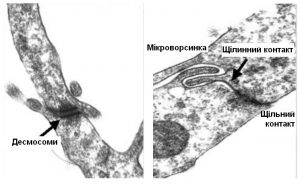
Характерно, що всі клітини трофектодерми – це розплющені клітини. На них діє тиск рідини в порожнині й тому вони мають пласку форму. Вони міцно з’єднані між собою щільними контактами, замикаючими мікроворсинками, десмосомами (Рис.3). Але на полюсі віддаленому від ВКМ з’являється групка клітин опуклої форми, яких назвали «руйнаторами блискучої оболонки». Мені не вдається придумати такого ж крутого терміна як англійське «zona breaker», можливо «клітини-розривачі» або «клітини-відмички» (Рис.4). Завдяки трансмісійній електронній мікроскопії вдалось отримати зображення мікроструктури цих клітин і таки зрозуміти їх функцію.
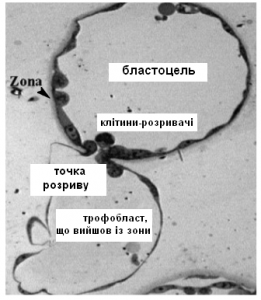
Виявилось, що ці клітини в місці контакту з оболонкою формують величезну кількість мікроворсинок (Рис.5, MV – мікроворсинки). Схожі мікроворсинки мають клітини тонкого кишківника для збільшення площі поверхні і всмоктування поживних речовин, а також клітини внутрішнього вуха і вушної завитки, завдяки ним ми чуємо звуки. Мікроворсинки всередині стабілізовані білком актином, а не мікротрубочками з тубуліну, як клітинні війки. Ці клітини мають інтенсивну секреторну активність. В їх цитоплазмі багато лізосом і вони вивільняють вміст числених секреторних гранул. Скоріш за все, ці гранули містять багато протеолітичних ферментів, що розщеплюють білки оболонки. Тому процес хетчингу почали вважати напівмеханічним, напівхімічним процесом.
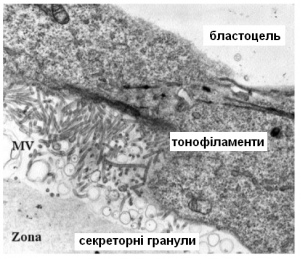
Після прориву ці ж клітини стабілізують дірку. Під мембраною клітини, що контактує з краєм Zona формується пояс тонофіламентів (Рис 6, TF – тонофіламенти) – ниток міцного білка кератину (так, саме того ж білка з якого складається волосся та нігті). Такі нитки утримують цей «сфінктер» відкритим. Тонофіламенти – активні учасники формування контактів між клітинами – десмосом. Такі контакти формуються в клітинах, на які постійно здійснюється механічний тиск: в шкірі, міокарді, є їх трохи в селезінці, печінці та нервових клітинах, ну й у трофектодермі їх багато. Це ніби як точкове зварювання клітин. Такі контакти дуже міцні, можливо десмосоми теж мають вивчати в Інституті зварювання Патона?
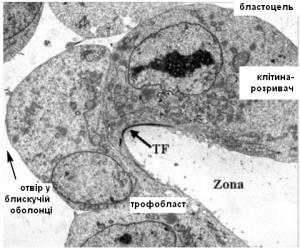
Досить цікавою є така полярність трофектодерми. Клітини-руйнатори оболонки віддалені від ВКМ. А от після хетчингу в процесі імплантації, проростання ембріона в шари ендометрія матки, там уже працюють клітини, що знаходяться ближче до ВКМ. Саме з них формується синцитієтрофобласт – оперативна група клітин, що виконуює інвазію і вростання в кровоносні судини матки.
Далі на ембріон ще чекає велика кількість пригод і контрольних точок розвитку, але хетчинг для людських ембріонів – це майже найголовніший крок у вільне і світле майбутнє. Переможці ембріогенезу отримують в подарунок додаткову порцію випробувань, а екзамени, співбесіди, відмова гарного хлопця – це все дрібниці в порівнянні з тією боротьбою за життя, яку ніхто з нас не пам’ятає.
Джерело
P.S. від редакції: щоб не пропустити новинок – підписуйтесь на наш канал у телеграмі t.me/my_science_ua
Дні науки

Наші проєкти

Щеплення Правдою



Обговорення
19 Вересня 2019, 12:32
А чому власне Зона пелюціда така жорстка? Що там у складі окрім білків? Що забезпечує жорсткість?
19 Вересня 2019, 13:55
Ну там не просто білки, а глікопротеїни, вони всі глікозильовані з купою вуглеводних залишків, які переплітаються, зшиваються між собою, ну і є певний відсоток глікозаміногліканів для “желєйки”)
21 Вересня 2019, 00:10
Дякую!
Напишіть відгук