Таємниця походження пожирачів вірусів
23 Березня 2017 | Молекулярно про Рак
Категорія: Біологія
Теги: віросвіт, вірофаги, вірусологія, гігантські віруси, евкаріогенез, еволюція, протисти, транспозони
Із загального курсу біології всім відомо, що віруси — молекулярні паразити клітинних форм життя: бактерій, архей та евкаріотів. Але неймовірно строкатий віросвіт не припиняє нас дивувати, і, як ми побачили в минулій публікації, самі віруси можуть ставати жертвами інших вірусів — так званих вірофагів (найбільш вивчені представники — Sputnik та Mavirus). Ці «молекулярні паразити в квадраті» уражають гігантські віруси одноклітинних евкаріотів (протистів), таких як Acanthamoeba polyphaga та Cafeteria roenbergensis. Діяльність вірофага заважає гігантському вірусу розмножуватися, що полегшує протікання епідемії в популяції протистів і сприяє виживанню більшої кількості одноклітинних організмів. Зі свого боку, протисти радо користуються такими анти-гігантовірусними властивостями вірофагів і навіть «дозволяють» останнім вибудовуватись у свій геном. Проте, така поведінка є нетиповою для ДНК вірусу: зазвичай він тримає свою ДНК окремо від клітинної. Інтеграція в клітинний геном — характерна риса іншого типу егоїстичних генетичних елементів під назвою транспозони. Більш того, вірофаг Mavirus має в своєму арсеналі гени, типові для транспозонів групи Maverick/Polinton, зокрема інтегразу (від англ. integrate — вбудовуватися), яку він використовує для потрапляння в клітинну ДНК. Цей факт прямо вказує на його близьку спорідненість із транспозонами зазначеної групи. Але тут виникає класична дилема курки і яйця: вірофаги дали початок траспозонам або навпаки?
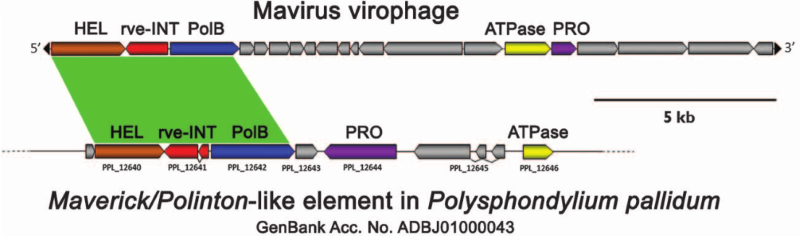
На думку Fischer, M.G. та Suttle, C.A., які вперше описали Mavirus в 2011 році, саме вірофаг перетворився на транспозон. В них було блискуче, хоча й вкрай спекулятивне на той час, пояснення своєї гіпотези. Наявність інтегрази свідчила про теоретичну здатність Mavirus вбудовуватися в геном джгутиконосця C. roenbergensis. Але водночас самій клітині вигідно «мати при собі» вірофаг, який протидіє явній загрозі з боку гігантського вірусу (Cafeteria roenbergensis virus, CroV). Така кооперація між клітиною та вірофагом створює передумови до накопичення вірофагів у геномі клітини. Проте з часом, особливо за відсутності тиску з боку гігантського вірусу, частина інтегрованих послідовностей «псується» за рахунок нейтральної молекулярної еволюції, внаслідок чого вони втрачають гени, важливі для реактивації, і таким чином перестають бути вірусами. Саме таку модель перетворення вірусу на транспозон запропонували автори. Результати подальших експериментів, опублікованих нещодавно в журналі Nature, чітко доводили той факт, що Mavirus дійсно вбудовується в геном C. roenbergensis. Також їхні дані з упевненістю свідчили на користь того, що вірофаг, навіть в інтегрований формі, перешкоджає поширенню CroV в популяції клітин. Тож, здавалося б, модель «вірофаг -> Polinton» виглядала доволі переконливою. Однак глобальне бачення еволюційної історії траспозонів явно не узгоджувалося із таким сценарієм.
Вперше на це звернули увагу Yutin, N., Raoult, D. та Koonin, E. В 2013 році вони публікують своє бачення походження вірофагів засноване на результатах молекулярно-філогенетичного аналізу консервативних протеїнів
Тут — невеличкий відступ для швидкого пояснення, шо малось на увазі. Консервативними є протеїни, які конче необхідні для життєдіяльності та розмноження вірусів, зокрема ті, що відповідають за утворення оболонок (великий і малий капсидні білки та цистеїнова протеаза, яка їх активує), копіювання ДНК (ДНК полімераза) та пакуванні ДНК всередину віріону (АТФаза), див. Рис. 1 вище. Оскільки ці компоненти незамінні, вони успадковуються в поколіннях вірусів із мінімальними змінами амінокислотного складу. Амінокислотні послідовності білків з різних представників вірофагів можна напряму порівняти на схожість методом так званого вирівнювання (приблизно так само як би ви вирівнювали два рядки цифр — виграшну комбінацію і вказану вами у лотерейному білеті –, щоб порахувати скільки цифр ви вгадали і на який виграш можете претендувати): чим більше різняться послідовності тим далі споріднені відповідні віруси (як от троюрідні брати), і навпаки, чим менша різниця, тим віруси «рідніші». Далі, знаючи усі відстані між вірусами, будується «родовід» (так зване філогенетичне дерево), з якого можна з’ясувати які групи виокремилися раніше, а які — еволюційно молодші
Отже, за даними дослідників, усі вірофаги розподілялися на дві еволюційні гілки, одна з яких представлена Mavirus, інша — Sputnik. Але обидві ці гілки сходилися в одній точці. Це означало, що згадані вірофаги пішли від спільного предка. Такий розподіл підтверджувався аналізом усіх консервативних протеїнів, окрім одного — ДНК полімерази. Виявилося, що вони взагалі належать до різних класів: у Sputnik — схожа на бактеріальну полімеразу першого типу (TVpol), тоді як у Mavirus — це полімераза типу Б (pPolB). Як ми побачимо далі, саме ця маленька зачіпка виведе нас на таємничого спільного предка вірофагів.
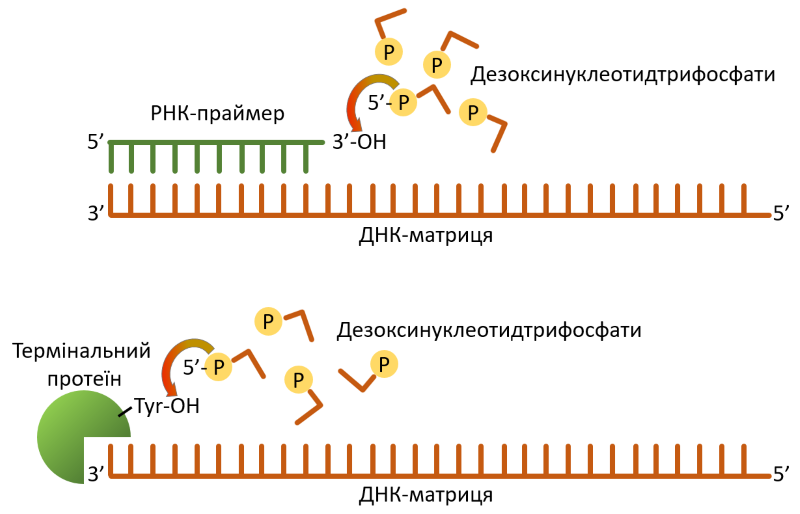
Полімераза типу Б є унікальною в плані механізму початку синтезу ДНК (так званого праймування, Рис. 2). В той час як звичайні полімерази потребують РНК затравку для старту матрице-залежного копіювання ДНК, pPolB додає перший дезоксирибонуклетидтрифосфат до –OH-групи залишку тирозину, серину або треоніну на термінальному протеїні, відтак, новоутворена ДНК буде ковалентно зв’язана із зазначеним протеїном. Така стратегія праймування не використовується жодним клітинним організмом — лише вірусами і … транспозонами групи Polinton. «Збіг? — навряд чи!» — подумали вчені, і вирішили перевірити в яких відносинах перебувають вірофаги та полінтони. На щастя, в обох згаданих егоїстичних генетичних елементах знайшлися спільні консервативні протеїни, а саме, цистеїнова протеаза та пакуюча АТФаза, що уможливило пряме порівняння і побудову загального «родоводу» за кожним із зазначених генів.
Виявилося, що на обох філогенетичних деревах (як за протеазою, так і за АТФазою) вірофаги є лише невеличким бічним пагоном в гущавині розмаїтої еволюційної гілки полінтонів. Така схема розташування (ще й підтримана відразу двома консервативними протеїнами) однозначно вказувала на те, що спільним предком всіх вірофагів був транспозон групи Polinton. Такий самий результат був отриманий за підсумком аналізу pPolB вірофагу Mavirus (Рис. 3). На жаль, Sputnik принципово неможливо всадовити на одне дерево разом із полінтонами за послідовністю полімерази, оскільки вона належить до іншого класу — TVpol. Дослідники пояснюють це заміщенням гена полімерази у предкового Sputnik-вірофага шляхом випадкової рекомбінації із іншим невідомим вірусом. Взагалі, така мобільність генів є характерною особливістю світу вірусів і транспозонів.
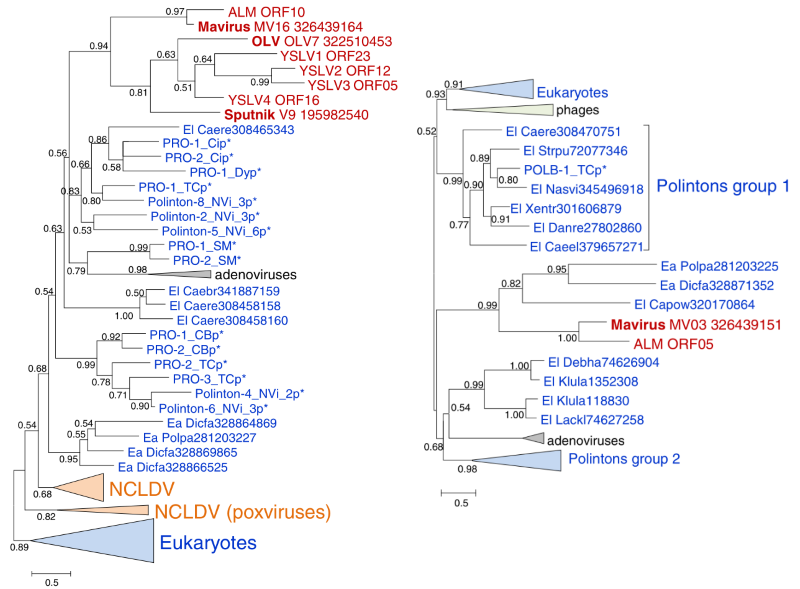
За рік після публікації результатів філогенетичного аналізу вірофагів було зроблено ще одне цікаве відкриття: в геномах гідри (Hydra magnipapillata) та рибки Danio rerio знайшли полінтони, які мали гени, схожі на капсидні білки ДНК вірусів. З цього виходило, що в минулому Polinton-траспозони були справжніми вірусами, а, можливо, деякі з них ще не втратили здатності до утворення активних вірусних часток, хоча прямих доказів останнього твердження дослідники не мали. Проте, залишалося принципове питання: якщо вірофаги не були предками полінтонів, тоді як же виглядав той вірус, який зрештою став полінтоном?
Ймовірний еволюційний сценарій, що відповідав на поставлене запитання, був викладений в оглядовій статті Nature Reviews Microbiology авторства Krupovic, M. та Koonin, E., 2015 року (Рис. 4). Їхня гіпотеза передбачала древнє походження полінтонів, що сягає моменту виникнення самих евкаріотів. Такого висновку дослідники дійшли на підставі поширеності полінтонів серед всіх основних супергруп евкаріотів (за виключенням рослин). Відтак, Krupovic, M. та Koonin, E. висувають сміливе припущення, що предком згадуваних транспозонів може бути вірус бактерій — бактеріофаг з родини Tectiviridae, оскільки він поділяє із полінтонами аж чотири гомологічні гени: великий і малий капсиди, пакуючу АТФазу, а також згадувану вище унікальну полімеразу типу Б. Вони пояснюють, що бактеріофаг міг потрапити до евкаріотичної клітини разом із альфа-протеобактерією — предком сучасної мітохондрії. Відомо, що процес встановлення ендосимбіозу між предком всіх евкаріотів та майбутньою мітохондрією супроводжувався активним перенесенням генів в напрямку від мітохондрії до евкаріта — так званий ендосимбіотичний генний трансфер. Цей феномен, як вважали вчені, відповідальний за появу послідовності ДНК бактеріофагу в евкаріотичному геномі.
Цікаво, що сучасні представники родини Tectiviridae нездатні до інтеграції в бактеріальний геном. Тому логічно припустити, що предок полінтонів також не мав такої можливості, а отримав її лише пізніше за рахунок рекомбінації із іншими егоїстичними генетичними елементам, що були присутні у тодішнього предка евкаріотів. Зокрема, полінтоніва інтеграза (ензим, що відповідає за вбудовування в ДНК клітини-хазяїна) за даними усе того ж філогенетичного аналізу походить від ретротранспозонів — окремого класу мобільних генетичних елементів, здатних до зворотної транскрипції. Саме така гібридна стратегія виживання дозволила напів-вірусу напів-транспозону досягнути неабиякого еволюційного успіху, що виражався у повсюдному поширенні серед евкаріотів. Надалі частина цих успішних гібридів обрала шлях втрати своєї вірусної частини і перетворилася на справжні транспозони. Інші навпаки — перестали покладатися на можливість інтеграції в чужинну ДНК і стали типовими вірусами (на думку авторів, саме так вчинили предки сучасних аденовірусів). А герої нашої розповіді — вірофаги — так і не змогли відмовити собі у розкоші вести подвійне життя, і обрали шлях паразитування на гігантських вірусах.
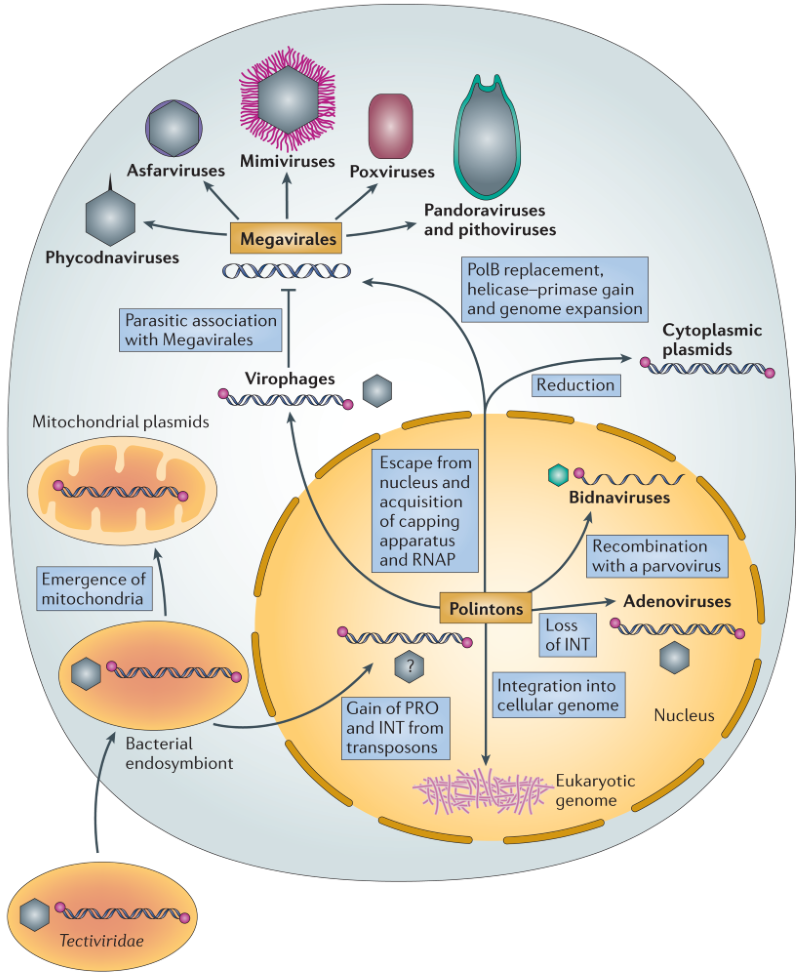
До останнього часу загально прийнятою тезою було та, що вірус принципово відрізняється від мобільного генетичного елементу здатністю утворювати віріони, побудовані з капсидних білків. Але головний висновок з усього цього наукового детективу — насправді немає чіткої межі між вірусами і транспозонами. Причина такої перемішаності не тільки в тому, що ці два класи генетичних паразитів обмінюються генами як привітальними листівками, але й в тому, що їхні еволюційні історії тісно переплетені. Найбільш очевидним це стало після детального вивчення походження вірофагів — від вірусів бактерій до евкаріотичних транспозонів і, зрештою, назад до типових вірусів, які зберегли можливість вбудовуватися в геном одноклітинних істот. Звичайно, не слід забувати, що еволюційні сценарії будуються виходячи із наявних даних і доступних методів їх обробки, тож є лише гіпотетичними, а в деяких випадках — навіть спекулятивними. Однак, слідкувати за науковими дебатами і грою аргументів іноді буває доволі цікаво.
Даниїл Грибчук для Моєї Науки
- Krupovic, Mart; Koonin, Eugene V. (2014). Polintons: a hotbed of eukaryotic virus, transposon and plasmid evolution. Nature Reviews Microbiology 13 (2). с. 105–115. doi:10.1038/nrmicro3389.
- Yutin, Natalya; Raoult, Didier; Koonin, Eugene V (2013). Virophages, polintons, and transpovirons: a complex evolutionary network of diverse selfish genetic elements with different reproduction strategies. Virology Journal 10 (1). с. 158. doi:10.1186/1743-422X-10-158.
Дні науки

Наші проєкти

Щеплення Правдою



Обговорення
6 Жовтня 2017, 10:51
а я вже думав, що малюнків нормальної якості вже і не побачу 🙂
ЗІ: дякую
Напишіть відгук